Familia Trochodendraceae (= Ordo Trochodendrales)
Sono tutte tropicali. Possiedono fiori ermafroditi con perianzio ridotto.
martedì 30 dicembre 2008
Familia Platanaceae

Familia Platanaceae
Il genere Platanus possiede fiori unisessuali estremamente ridotti, riuniti in infiorescenze da cui derivano le infruttescenze. L’ovario è coricarpico.
Secondo alcuni autori i fiori estremamente ridotti sono considerati un carattere evoluto che accomuna le platanacee alle fagacee, mentre secondo le analisi cladistiche dell’APG sono un carattere primitivo che accomuna le platanacee alle angiosperme più primitive.
Clade delle Eudicotiledoneae
Clade delle Eudicotiledoneae
I caratteri apomorfici che identificano le eudicotiledoni sono: fiori tetra o pentameri, eteroclamidati e gamopetali, polline triaperturato.
Gli ordini contenuti in questo clade si riuniscono in gruppi, in base al grado evolutivo:
Triaperturatae = Eudicotiledoneae
Eudicotiledoneae arcaiche
Eudicotiledoneae con caratteri arcaici ed evoluti
Eudicotiledoneae superiori
I caratteri apomorfici che identificano le eudicotiledoni sono: fiori tetra o pentameri, eteroclamidati e gamopetali, polline triaperturato.
Gli ordini contenuti in questo clade si riuniscono in gruppi, in base al grado evolutivo:
Triaperturatae = Eudicotiledoneae
Eudicotiledoneae arcaiche
Eudicotiledoneae con caratteri arcaici ed evoluti
Eudicotiledoneae superiori
Ordo Proteales

Ordo Proteales
Le proteali sono l’ordine più primitivo. Comprendono differenti famiglie Platanaceae, Trochodendraceae, Nelumbonaceae e le Proteaceae (tutte esotiche ed utilizzate come piante ornamentali).
lunedì 29 dicembre 2008
Ordo Laurales
Ordo Laurales
Gruppo di alberi con caratteri primitivi. Possiedono un ovario semiinfero e molte cellule secretrici con oli essenziali. Nella famiglia delle Lauraceae sono compresi Laurus nobilis (alloro, dioico), Cinnamomum canphora (canfora, estratta dal legno per sublimazione), Cinnamomum zeylanicum (cannella), Persea gratissima (avocado).
Gruppo di alberi con caratteri primitivi. Possiedono un ovario semiinfero e molte cellule secretrici con oli essenziali. Nella famiglia delle Lauraceae sono compresi Laurus nobilis (alloro, dioico), Cinnamomum canphora (canfora, estratta dal legno per sublimazione), Cinnamomum zeylanicum (cannella), Persea gratissima (avocado).
domenica 28 dicembre 2008
Familia Myristicaceae

Familia Myristicaceae
Comprende Myristica fragrans, la noce moscata. Possiede frutti avvolti da un arillo carnoso, rosso, con endosperma ruminato ed intersecato dalla nucella.
Familia Magnoliaceae

Familia Magnoliaceae
Comprende Magnolia grandiflora, Liriodendron papirifera. Sono alberi imponenti con caratteri molto primitivi.
Familia Degeneriaceae

Familia Degeneriaceae
Nelle diverse specie del genere Degeneria è visibile l’evoluzione degli stami. L’ovario è primitivo.
Ordo Magnoliales
Ordo Magnoliales
Gruppo di alberi con caratteri molto primitivi. I fiori sono grandi e con organizzazione spiralata, l’ovario è supero e apocarpico.
Gruppo di alberi con caratteri molto primitivi. I fiori sono grandi e con organizzazione spiralata, l’ovario è supero e apocarpico.
Gruppo dei paleoalberi
Gruppo dei paleoalberi
Insieme alle paleoerbe costituiscono il gruppo basale più probabile dalle quali sono derivate le altre angiosperme. Alcuni autori pensano che le angiosperme primitive avessero un portamento ad alberello simile a quello di alcuni paleoalberi.
Ordo Nympheales
Ordo Nympheales
Questo ordine non è stato individuato con certezza, comprendono la famiglia delle Nympheaceae (genere Nymphaea, Nuphar luteum). Non hanno cellule secretrici, hanno sepali petaloidei e presentano tutta la serie di passaggio tra petali e stami. Spesso i cotiledoni sono fusi (sincotilia).
Per Cronquist sono l’anello di congiunzione tra i paleoalberi (o magnolide) e le eudicotiledoni.
Ordo Piperales

Ordo Piperales
Alcuni caratteri le avvicinano alle monocotiledoni, poiché possiedono fiori con stami e perianzio trimeri, il fusto con atactostele ma con fasci aperti (vi è quindi un cambio tra floema e xilema).
I caratteri tipici dell’ordine compredono: fiori apetali e cellule secretrici.
Compredono la famiglia delle Aristolochiaceae e delle Piperaceae (Piper nigrum, i cui frutti immaturi essicati danno il pepe nero, i frutti maturi e sbucciati danno il pepe bianco).
sabato 27 dicembre 2008
Ordo Ceratophyllales

Ordo Ceratophyllales
Ceratophyllum è il genere forse più vicino all’angiosperma originaria.
Gruppo delle paleoerbe
Gruppo delle paleoerbe
Gli ordini di paleoerbe erano prima inclusi nelle dicotiledoni. Oggi sono poste a parte ed accomunate dall’avere caratteri primitivi e un portamento erbaceo.
Gli ordini di paleoerbe erano prima inclusi nelle dicotiledoni. Oggi sono poste a parte ed accomunate dall’avere caratteri primitivi e un portamento erbaceo.
Clade delle Magnoliidae

Clade delle Magnoliidae
Le magnoliide comprendono ordini di dicotiledoni primitive, le paleoerbe e i paleoalberi. Possiedono diversi caratteri plesiomorfici comuni alle monocotiledoni (sopratutto con quelle più arcaiche) tra cui: pollini inaperturati, entomofilia (spesso cantarofilia), perianzio non specializzato (non si riconoscono calice e corolla), stami spesso imperfetti.
I caratteri tipici e apomorfi riguardano: le foglie che hanno nervature pennate, le trachee che sono imperfette o assenti. I fiori sono trimeri e/o a disposizione spiralata e il polline monoaperturato o derivato da questo.
venerdì 26 dicembre 2008
Paleontologia delle angiosperme
Paleontologia delle angiosperme
La maggior parte dei fossili di antiche angiosperme risale al Giurassico ed al Cretaceo, mentre nel Terziario si ritrovano già tutti i gruppi principali, arrivati sino ad oggi.
Tuttavia, il fossile più antico conosciuto è Sanmiguenia, del Triassico, che possedeva foglie grandi e parallelinervie ed infiorescenze maschili e femminili differenziate, alle quali sono stati attribuiti nomi diversi. I fossili, essendo spesso incompleti, non forniscono dati molto precisi.
Nei successivi periodi del Mesozoico, si ha una grande differenziazione, soprattutto nel Cretaceo, dove si ritrovano molti pollini fossili monoaperturati, mentre nel Terziario prevalgono i pollini triaperturati.
La relativa mancanza di fossili nelle prime fasi evolutive delle angiosperme, può essere dovuta al loro luogo di origine. Si ipotizza che l’evoluzione delle prime angiosperme, nel Triassico, sia avvenuta in zone montane in cui vi era scarsità di anidride carbonica e quindi dove non vi era molta probabilità di fossilizzazione. Alcuni autori pensano che il clima, in queste zone montane, fosse semiarido, mentre altri pensano fosse un clima stagionale. In seguito colonizzarono anche gli ambienti planiziali dove sarebbero state avvantaggiate dalle loro conquiste evolutive.
La maggior parte dei fossili di antiche angiosperme risale al Giurassico ed al Cretaceo, mentre nel Terziario si ritrovano già tutti i gruppi principali, arrivati sino ad oggi.
Tuttavia, il fossile più antico conosciuto è Sanmiguenia, del Triassico, che possedeva foglie grandi e parallelinervie ed infiorescenze maschili e femminili differenziate, alle quali sono stati attribuiti nomi diversi. I fossili, essendo spesso incompleti, non forniscono dati molto precisi.
Nei successivi periodi del Mesozoico, si ha una grande differenziazione, soprattutto nel Cretaceo, dove si ritrovano molti pollini fossili monoaperturati, mentre nel Terziario prevalgono i pollini triaperturati.
La relativa mancanza di fossili nelle prime fasi evolutive delle angiosperme, può essere dovuta al loro luogo di origine. Si ipotizza che l’evoluzione delle prime angiosperme, nel Triassico, sia avvenuta in zone montane in cui vi era scarsità di anidride carbonica e quindi dove non vi era molta probabilità di fossilizzazione. Alcuni autori pensano che il clima, in queste zone montane, fosse semiarido, mentre altri pensano fosse un clima stagionale. In seguito colonizzarono anche gli ambienti planiziali dove sarebbero state avvantaggiate dalle loro conquiste evolutive.
giovedì 25 dicembre 2008

Diversi sistemi di classificazione delle angiosperme
Diversi sistemi di classificazione delle angiosperme
Un primo tentativo di costruire una classificazione naturale viene effettuato dalla scuola germanica di Engler e Wettstein. Per Engler e, successivamente, Wettstein l’idea portante è che le forme semplici siano da interpretare come primitive e le forme complesse come evolute. Essi ritengono, inoltre, più primitive le piante con fiori grandi isolati e più evolute quelle con fiori piccoli riuniti in infiorescenze che simulano grandi fiori. Considerano tra i caratteri importanti la posizione dell’ovario (che da supero, diviene infero), il concrescimento dei carpelli, etc.
Dal punto di vista filogenetico Engler e Wettstein non tengono conto della possibile riduzione e atrofia di parti fiorali nel corso dell’evoluzione.
Sistema di classificazione di Engler (1897)
Angiosperme
Dicotiledoni
Archiclamidee
Monoclamidee
Dialipetale
Metaclamidee
Monocotiledoni
Fino ad oltre il 1950 il sistema seguito è quello della scuola germanica. Alla fine dell’800 nasce anche la scuola anglosassone, cominciata da Bessey. Egli costruisce una prima classificazione filogenetica che tiene conto delle riduzioni e delle atrofie che possono semplificare secondariamente caratteri complessi.
Per esempio: per la scuola germanica le “amentifere” (piante con amenti) sono considerate le più primitive poiché anemofile, mentre per la scuola anglosassone le Magnoliaceae sono le più primitiva a causa del loro fiore. Secondo la scuola anglosassone le “amentifere” hanno sviluppato secondariamente l’anemofilia e l’unisessualità.
È proprio dalla logica seguita dalla scuola anglosassone che derivano le classificazioni moderne.
Le classificazioni moderne si basano sempre su una vasta gamma di caratteri, tra i quali vengono presi in considerazione, oltre a quelli morfologico-anatomici, sempre più caratteri individuati con le analisi sierologiche, la determinazione delle sequenze degli amminoacidi per le proteine o dei nucleotidi degli acidi nucleici, lo studio della biosintesi di metabolici secondari e le osservazioni ultrastrutturali.
Takhtajan (1967-1969) mette in evidenza alcuni fatti che emergono dai dati disponibili:
La probabile origine monofiletica delle angiosperme.
Anche se non si conoscono fossili sicuri prima del Cretaceo, probabilmente, sono da interpretarsi come angiosperme dei fossili del Giurassico e forse della fine del Triassico.
Il luogo di origine delle angiosperme va ricercato dove esistono numerose famiglie e generi con caratteri di primitività, cioè in Asia sudorientale, tra l’Assam e le isole Fiji (parti dell’originale Gondwana).
Cronquist (1968) delinea uno schema fondamentale simile a quello di Takhtajan, col quale è stato in parte integrato. Lo schema tassonomico seguito dalla maggior parte dei testi didattici è quello di Takhtajan e Cronquist, eventualmente con qualche modifica.
Takhtajan e Cronquist costruiscono una classificazione filogenetica molto utilizzata poiché ancora caratterizzata da una certa praticità.
Un nuovo schema di classificazione, che tiene conto in particolare dell’analisi cladistica dei dati morfologici e biomolecolari, è stato proposto da un gruppo di ricercatori, l’APG (Angiosperm Phylogeny Group) nel 1998. Naturalmente anch’esso potrà essere modificato in seguito all’apporto di nuovi dati.
str
La tendenza attuale è nel distinguere due principali tipi di classificazione a seconda dell’uso a cui sono destinate: classificazioni pratiche (utilizzate nelle flore) e classificazioni moderne filogenetiche (utilizzate nell’analisi filogenetica).
Tappe evolutive dell'origine delle Magnoliophyina
Tappe evolutive dell'origine delle Magnoliophyina
Pteridofite primitive: felci isosporee con gametofiti liberi.
Pteridofite derivate: felci eterosporee con gametofiti germinanti dentro le spore.
Conservazione della spora nello sporangio e sporocarpi protettivi
Pteridosperme: Formazione dell'ovulo con gametofito femminile racchiuso nel macrosporangio, che diviene la nucella di nutrimento, gametofiti maschili liberi (polline) dotati di tubetto pollinico e spermatozoidi flagellati.
Angiosperme: chiusura degli ovuli nell'ovario e stimma per catturare il polline, micropilo nascosto e raggiunto solo dai nuclei spermatici. Vi è una tendenza nel prolungare il percorso del tubetto pollinico per permettere una rigida selezione dei nuclei spermatici ed evitare l'autogamia.
Pteridofite primitive: felci isosporee con gametofiti liberi.
Pteridofite derivate: felci eterosporee con gametofiti germinanti dentro le spore.

Conservazione della spora nello sporangio e sporocarpi protettivi
Pteridosperme: Formazione dell'ovulo con gametofito femminile racchiuso nel macrosporangio, che diviene la nucella di nutrimento, gametofiti maschili liberi (polline) dotati di tubetto pollinico e spermatozoidi flagellati.
Angiosperme: chiusura degli ovuli nell'ovario e stimma per catturare il polline, micropilo nascosto e raggiunto solo dai nuclei spermatici. Vi è una tendenza nel prolungare il percorso del tubetto pollinico per permettere una rigida selezione dei nuclei spermatici ed evitare l'autogamia.
mercoledì 24 dicembre 2008
dispersione idrocora
Dispersione idrocora: il frutto possiede delle strutture per il galleggiamento, grazie alle quali colonizza diverse aree. Ne è un esempio il cocco, che galleggia sull’acqua rimanendo però protetto al suo interno da un involucro lignificato.
dispersione autocora
Dispersione autocora: la pianta stessa spara i semi lontano. Per esempio i legumi che si aprono bruscamente scagliando i semi (in Ecballium elaterium, il cocomero asinino, il frutto scoppia).
Dispersione anemocora
Dispersione anemocora: questo tipo di dispersione viene attuata tramite pappi o ali che permettono al seme di essere portato lontano dalla pianta madre.
dispersione zoocora
Dispersione zoocora: questo tipo di dispersione avviene tramite animali, o l’uomo (dispersione antropocora). Le specie che sfruttano questo tipo di dispersione hanno spesso frutti carnosi che vengono ingeriti dagli animali, mentre i semi sono espulsi intatti. Se invece possiedono frutti secchi, questi hanno appendici con cui si impigliano nel pelo o nelle piume. Un tipo particolare di dispersione zoocora è quella mirmecocora, dove i semi vengono trasportati dalle formiche. I semi possiedono escrescenze carnose (strofiolo e caruncola) che vengono mangiate dalle formiche, senza che si cibino anche del seme, che viene disperso.
martedì 23 dicembre 2008
semi ed embrione
Semi ed embrione
Dopo la fecondazione dell’oosfera si ha la formazione del seme a partire dai tegumenti degli ovuli. Nel seme, a maturazione, possiedono la testa e i tegumenti. I tegumenti del seme sono 2 o 1 a seconda dei tegumenti dell’ovulo da cui deriva. Vi sono poi altre strutture accessorie: lo strofiolo (un’escrescenza carnosa del funicolo dell’ovulo), la caruncola (un’escrescenza della zona micropilare), l’ilo (punto in cui si attacca il funicolo).
Il seme contiene anche i tessuti di riserva derivanti dalla seconda fecondazione dei nuclei polari, che formano l’endosperma secondario 3n. Questa sostanza di riserva può essere cellularizzata o meno, la nucella rimanente prende il nome di perisperma. In alcune dicotiledoni l’endosperma viene riassorbito ed il perisperma viene contenuto nei cotiledoni.
I semi si distinguono in base al tipo di nutrimento che contengono:
Semi albuminosi: possiedono l’endosperma secondario come sostanza di riserva.
Semi escalbuminosi: possiedono il perisperma nei cotiledoni come sostanza di riserva.
Vi sono poi alcuni casi particolari:
Semi di ricino: sono semi albuminosi con granuli di aleurone e gocce d’olio.
Semi di agrostena: l’embrione è avvolto all’endosperma.
Semi di lino: il tegumento ha funzione di riserva.
All’interno del seme è contenuto l’embrione 2n, derivante dalla fecondazione dell’oosfera, esso è costituito da:
1 cellula basale
Cellule del sospensore
1 cellula apicale
La cellula apicale si divide e differenzia in regioni che a loro volta si differenzieranno nella pianta adulta:
Radichetta, posta nella parte basale.
Asse ipocotile
Cotiledoni, al centro dell’embrione.
Asse epicotile
Plumula, all’apice dell’embrione.
Inizialmente l’embrione è ripiegato su se stesso nel seme, poi grazie alle divisioni cellulari l’asse epicotile si rivolge verso l’alto e l’embrione si dispiega.
Dopo la fecondazione dell’oosfera si ha la formazione del seme a partire dai tegumenti degli ovuli. Nel seme, a maturazione, possiedono la testa e i tegumenti. I tegumenti del seme sono 2 o 1 a seconda dei tegumenti dell’ovulo da cui deriva. Vi sono poi altre strutture accessorie: lo strofiolo (un’escrescenza carnosa del funicolo dell’ovulo), la caruncola (un’escrescenza della zona micropilare), l’ilo (punto in cui si attacca il funicolo).
Il seme contiene anche i tessuti di riserva derivanti dalla seconda fecondazione dei nuclei polari, che formano l’endosperma secondario 3n. Questa sostanza di riserva può essere cellularizzata o meno, la nucella rimanente prende il nome di perisperma. In alcune dicotiledoni l’endosperma viene riassorbito ed il perisperma viene contenuto nei cotiledoni.
I semi si distinguono in base al tipo di nutrimento che contengono:
Semi albuminosi: possiedono l’endosperma secondario come sostanza di riserva.
Semi escalbuminosi: possiedono il perisperma nei cotiledoni come sostanza di riserva.
Vi sono poi alcuni casi particolari:
Semi di ricino: sono semi albuminosi con granuli di aleurone e gocce d’olio.
Semi di agrostena: l’embrione è avvolto all’endosperma.
Semi di lino: il tegumento ha funzione di riserva.
All’interno del seme è contenuto l’embrione 2n, derivante dalla fecondazione dell’oosfera, esso è costituito da:
1 cellula basale
Cellule del sospensore
1 cellula apicale
La cellula apicale si divide e differenzia in regioni che a loro volta si differenzieranno nella pianta adulta:
Radichetta, posta nella parte basale.
Asse ipocotile
Cotiledoni, al centro dell’embrione.
Asse epicotile
Plumula, all’apice dell’embrione.
Inizialmente l’embrione è ripiegato su se stesso nel seme, poi grazie alle divisioni cellulari l’asse epicotile si rivolge verso l’alto e l’embrione si dispiega.
lunedì 22 dicembre 2008


Siliqua e siliquetta, possiedono 2 valve con una lamina centrale (replum), un falso setto (Capsella, Cardamine).

Siliqua e siliquetta, possiedono 2 valve con una lamina centrale (replum), un falso setto (Capsella, Cardamine).

frutti
Frutti
Il frutto deriva da parti del fiore o da fiori interi o da infiorescenze. Racchiude i semi fino alla loro maturazione e ne favorisce la diffusione, infatti forma e funzione dipendono essenzialmente dalla modalità di diffusione. Generalmente si distinguono 3 tegumenti del frutto che derivano dalla parete dell’ovario:
- Pericarpo, unistratificato.
- Mesocarpo, pluristrtificato.
- Endocarpo, unistratificato.
Questi tre tegumenti assumono consistenza differente nelle differenti specie, generalmente distinguibili in frutti secchi e frutti carnosi.
Nei frutti secchi tutte le parti del frutto maturo sono secche (cellule morte).
Nei frutti carnosi pericarpo, endocarpo e soprattutto mesocarpo sono carnosi (cellule vive) a maturazione. La polpa è costituita dall’endocarpo che si sviluppa verso l’interno e tra i semi (agrumi e banane).
I frutti si possono essere:
- Frutti semplici: frutti singoli derivanti da un fiore.
- Frutti composti o infruttescenze: derivanti da un fiore.
- Frutti sinantocarpici: derivanti da infiorescenze.
I frutti si dividono anche sulla base dell’emissione dei semi:
- Frutti deiscenti: si aprono liberando i semi.
- Frutti indeiscenti.
- Frutti dirompenti.
Il frutto deriva da parti del fiore o da fiori interi o da infiorescenze. Racchiude i semi fino alla loro maturazione e ne favorisce la diffusione, infatti forma e funzione dipendono essenzialmente dalla modalità di diffusione. Generalmente si distinguono 3 tegumenti del frutto che derivano dalla parete dell’ovario:
- Pericarpo, unistratificato.
- Mesocarpo, pluristrtificato.
- Endocarpo, unistratificato.
Questi tre tegumenti assumono consistenza differente nelle differenti specie, generalmente distinguibili in frutti secchi e frutti carnosi.
Nei frutti secchi tutte le parti del frutto maturo sono secche (cellule morte).
Nei frutti carnosi pericarpo, endocarpo e soprattutto mesocarpo sono carnosi (cellule vive) a maturazione. La polpa è costituita dall’endocarpo che si sviluppa verso l’interno e tra i semi (agrumi e banane).
I frutti si possono essere:
- Frutti semplici: frutti singoli derivanti da un fiore.
- Frutti composti o infruttescenze: derivanti da un fiore.
- Frutti sinantocarpici: derivanti da infiorescenze.
I frutti si dividono anche sulla base dell’emissione dei semi:
- Frutti deiscenti: si aprono liberando i semi.
- Frutti indeiscenti.
- Frutti dirompenti.
Rischio di autogamia
Rischio di autogamia
L’autogamia è generalmente evitata dagli organismi ermafroditi per non abbassare la variabilità genetica della generazione successiva, in modo da garantirne il successo adattativi.
Per questo motivo, nel corso dell’evoluzione si sono sviluppate tecniche per impedire l’autogamia:
Proterandria: sviluppo precoce degli elementi sessuali maschili in un fiore ermafrodito.
Proteroginia: sviluppo precoce degli elementi sessuali femminili in un fiore ermafrodito.
Riconoscimento di parentela a livello stigmatico: lo stigma riconosce se il polline possiede lo stesso patrimonio genetico e gli impedisce di germinare, oppure germina ma il tubetto pollinico non penetra.
Barriere dopo la germinazione: il polline germina sullo stigma ma incontra barriere alla fecondazione.
Aborto: gli embrioni generati per autogamia vengono abortiti. Questo meccanismo è raro poiché richiede un alto dispendio energetico.
Talvolta l’autogamia è favorita. Questo accade quando l’ambiente impedisce l’impollinazione da parte di altri individui. Anche a questo scopo si sono evoluti meccanismi appositi:
Fiori autogami facoltativi: ricorrono all’autogamia solo se non avviene l’allogamia.
Fiori apomittici: si riproducono per apomissia, cioè quando un’oosfera si sviluppa in un embrione senza essere fecondata.
Fiori cleistogami: fiori che si autoimpollinano prima ancora di aprirsi.
L’autogamia è generalmente evitata dagli organismi ermafroditi per non abbassare la variabilità genetica della generazione successiva, in modo da garantirne il successo adattativi.
Per questo motivo, nel corso dell’evoluzione si sono sviluppate tecniche per impedire l’autogamia:
Proterandria: sviluppo precoce degli elementi sessuali maschili in un fiore ermafrodito.
Proteroginia: sviluppo precoce degli elementi sessuali femminili in un fiore ermafrodito.
Riconoscimento di parentela a livello stigmatico: lo stigma riconosce se il polline possiede lo stesso patrimonio genetico e gli impedisce di germinare, oppure germina ma il tubetto pollinico non penetra.
Barriere dopo la germinazione: il polline germina sullo stigma ma incontra barriere alla fecondazione.
Aborto: gli embrioni generati per autogamia vengono abortiti. Questo meccanismo è raro poiché richiede un alto dispendio energetico.
Talvolta l’autogamia è favorita. Questo accade quando l’ambiente impedisce l’impollinazione da parte di altri individui. Anche a questo scopo si sono evoluti meccanismi appositi:
Fiori autogami facoltativi: ricorrono all’autogamia solo se non avviene l’allogamia.
Fiori apomittici: si riproducono per apomissia, cioè quando un’oosfera si sviluppa in un embrione senza essere fecondata.
Fiori cleistogami: fiori che si autoimpollinano prima ancora di aprirsi.
domenica 21 dicembre 2008
Impollinazione
Impollinazione
Si pensa che l’impollinazione entomofila sia la più primitiva, mentre l’anemofilia sia secondaria e legata a particolari ambienti (al contrario delle gimnosperme). Anche oggi l’impollinazione entomofila è la più importante. Inizialmente gli insetti impollinatori (o pronubi) erano coleotteri che con il loro apparato masticatore aggredivano sia il polline che l’ovario. In questo modo le antiche angiosperme dovevano produrre una grande quantità di polline e proteggere l’ovario. Perciò si assiste ad un’evoluzione parallela di piante e insetti, fino ad avere rapporti specie specifici (infeudazione):
Coleotteri: impollinano fiori grandi e robusti.
Imenotteri: impollinano fiori con struttura dorsoventrale (di appoggio), corolla papilionacea o labiata.
Lepidotteri: impollinano fiori con corolle tubolari e petali speronati (in Madagascar, un’orchidea ha uno sperone di 32 cm!).
Per attirare gli insetti impollinatori si sono evoluti diversi accorgimenti per rendere i fiori più attraenti. Strutture secernenti il nettare, i nettari, con diverse derivazioni:
Nettari da stami.
Nettari dal ricettacolo.
Nettari dal perianzio.
Strutture colorate, secondo maculazioni o disegni che imitano le femmine corrispondenti dei maschi dell’insetto impollinatore. Strutture secernenti sostanze odorose, o profumate o con odori particolari (per esempio la Rafflesia arnoldi odora di carne putrescente per attirare ditteri). In casi particolari (orchidee) vengono secrete sostanze molto simili a ferormoni corrispondenti per le specie impollintrici.
In alcuni gruppi si sono evoluti meccanismi complessi e strutture specializzate per attirare gli insetti pronubi:
Fiori a trappola: sono piante dotate di spadice, una struttura sterile che accoglie l’infiorescenza in una fossetta. L’infiorescenza possiede fiori femminili alla base e maschili in alto, sovrastati da un ciuffo di peli. Gli insetti impollinatori sono ditteri che rimangono intrappolati dai peli. Quando arrivano i fiori maschili arrivano a maturazione, e rilasciano il polline sui ditteri. In seguito i peli si riaprono e liberano i ditteri che vanno a fecondare altri fiori.
Orchidee Ophris: il labello simula l’addome dell’insetto impollinatore ed emette feromoni. In alcune specie vi è uno specchietto in cui le gli insetti si riflettono!
Fico: sviluppa infiorescenze di due tipi, con ovari a stilo lungo e a stilo corto. Il suo insetto impollinatore è la Blastophaga: essa penetra nelle infiorescenze con stilo corto e deposita le uova. Ne nascono larve che si cibano degli ovari ma ne escono come insetti perfetti riempiti di polline. Questi, possono incontrare infiorescenze con ovari a stilo lungo, dove non riescono a deporre uova, limitandosi a impollinarle.
Salvia: il fiore possiede un antera sterile e una fertile. L’insetto impollinatore spinge l’antera sterile così che quella fertile tocca il suo addome, lasciando il polline.
In alcuni casi si assiste ad un tipo di impollinazione zoofila particolare:
Impollinazione ornitofila, presente soprattutto in piante tropicali. In Sud America vi sono i colibrì, in Africa le nettarine.
Impollinazione chirotterofila, cioè con pipistrelli. Sono fiori grossi e tubulosi con stigmi pennacchio.
Nelle angiosperme l’anemofilia è considerata sempre secondaria rispetto all’entomofilia. La tendenza al ritorno all’anemofilia può essere vantaggioso per specie che costituiscono formazioni monofitiche (per esempio nelle faggete) e zone con pochi insetti.
La transizione dall’entomofilia è visibile anche in differenti specie di uno stesso genere, per esempio: Sanguisorba officinalis con impollinazione entomofila e S. minor con impollinazione anemofila.
Una famiglia particolarmente adattata all’anemofilia sono le graminacee: nelle infiorescenze ciascun fiore possiede lunghi stami che vengono sbattuti dal vento in modo da disperderne il polline.
Un ultimo caso di impollinazione è l’idrofilia: nelle piante con impollinazione idrofila i fiori sono poco vistosi ed il polline è riunito in catenelle allungate, che vengono trasportate dall’acqua fino agli ovari.
Si pensa che l’impollinazione entomofila sia la più primitiva, mentre l’anemofilia sia secondaria e legata a particolari ambienti (al contrario delle gimnosperme). Anche oggi l’impollinazione entomofila è la più importante. Inizialmente gli insetti impollinatori (o pronubi) erano coleotteri che con il loro apparato masticatore aggredivano sia il polline che l’ovario. In questo modo le antiche angiosperme dovevano produrre una grande quantità di polline e proteggere l’ovario. Perciò si assiste ad un’evoluzione parallela di piante e insetti, fino ad avere rapporti specie specifici (infeudazione):
Coleotteri: impollinano fiori grandi e robusti.
Imenotteri: impollinano fiori con struttura dorsoventrale (di appoggio), corolla papilionacea o labiata.
Lepidotteri: impollinano fiori con corolle tubolari e petali speronati (in Madagascar, un’orchidea ha uno sperone di 32 cm!).
Per attirare gli insetti impollinatori si sono evoluti diversi accorgimenti per rendere i fiori più attraenti. Strutture secernenti il nettare, i nettari, con diverse derivazioni:
Nettari da stami.
Nettari dal ricettacolo.
Nettari dal perianzio.
Strutture colorate, secondo maculazioni o disegni che imitano le femmine corrispondenti dei maschi dell’insetto impollinatore. Strutture secernenti sostanze odorose, o profumate o con odori particolari (per esempio la Rafflesia arnoldi odora di carne putrescente per attirare ditteri). In casi particolari (orchidee) vengono secrete sostanze molto simili a ferormoni corrispondenti per le specie impollintrici.
In alcuni gruppi si sono evoluti meccanismi complessi e strutture specializzate per attirare gli insetti pronubi:
Fiori a trappola: sono piante dotate di spadice, una struttura sterile che accoglie l’infiorescenza in una fossetta. L’infiorescenza possiede fiori femminili alla base e maschili in alto, sovrastati da un ciuffo di peli. Gli insetti impollinatori sono ditteri che rimangono intrappolati dai peli. Quando arrivano i fiori maschili arrivano a maturazione, e rilasciano il polline sui ditteri. In seguito i peli si riaprono e liberano i ditteri che vanno a fecondare altri fiori.
Orchidee Ophris: il labello simula l’addome dell’insetto impollinatore ed emette feromoni. In alcune specie vi è uno specchietto in cui le gli insetti si riflettono!
Fico: sviluppa infiorescenze di due tipi, con ovari a stilo lungo e a stilo corto. Il suo insetto impollinatore è la Blastophaga: essa penetra nelle infiorescenze con stilo corto e deposita le uova. Ne nascono larve che si cibano degli ovari ma ne escono come insetti perfetti riempiti di polline. Questi, possono incontrare infiorescenze con ovari a stilo lungo, dove non riescono a deporre uova, limitandosi a impollinarle.
Salvia: il fiore possiede un antera sterile e una fertile. L’insetto impollinatore spinge l’antera sterile così che quella fertile tocca il suo addome, lasciando il polline.
In alcuni casi si assiste ad un tipo di impollinazione zoofila particolare:
Impollinazione ornitofila, presente soprattutto in piante tropicali. In Sud America vi sono i colibrì, in Africa le nettarine.
Impollinazione chirotterofila, cioè con pipistrelli. Sono fiori grossi e tubulosi con stigmi pennacchio.
Nelle angiosperme l’anemofilia è considerata sempre secondaria rispetto all’entomofilia. La tendenza al ritorno all’anemofilia può essere vantaggioso per specie che costituiscono formazioni monofitiche (per esempio nelle faggete) e zone con pochi insetti.
La transizione dall’entomofilia è visibile anche in differenti specie di uno stesso genere, per esempio: Sanguisorba officinalis con impollinazione entomofila e S. minor con impollinazione anemofila.
Una famiglia particolarmente adattata all’anemofilia sono le graminacee: nelle infiorescenze ciascun fiore possiede lunghi stami che vengono sbattuti dal vento in modo da disperderne il polline.
Un ultimo caso di impollinazione è l’idrofilia: nelle piante con impollinazione idrofila i fiori sono poco vistosi ed il polline è riunito in catenelle allungate, che vengono trasportate dall’acqua fino agli ovari.
androceo: coso dell'evoluzione
Nel corso dell’evoluzione si assiste ad una riduzione del numero di ovuli e ad un cambiamento della posizione degli ovuli nell’ovario (placentazione):
Placentazione assile.
Placentazione centrale.
Placentazione basale.
Inoltre l’ovulo può essere in diverse posizioni:
Atropo o ortotropo, cioè eretto, il tipo più primitivo.
Campilotropo, cioè orizzontale, il tipo intermedio.
Anatropo, cioè ripiegato, il tipo più comune ed evoluto.
Il gametofito femminile (sacco embrionale) non sviluppa mai archegoni. Si riduce a una struttura a 8 nuclei e 7 cellule. Degli 8 nuclei solo 3 si cellularizzano in posizione basale (cellule antipodali), altri 3 nuclei si cellularizzano nella parte apicale (cellule sinergidi) ai lati dell’oosfera. Gli ultimi 2 nuclei sono i nuclei polari liberi.
La doppia fecondazione avviene quando il tubetto pollinico penetra nelle cellule sinergidi: uno dei nuclei spermatici feconda l’oosfera, che originerà l’embrione 2n, mentre l’altro nucleo spematico si fonde con entrambi i nuclei polari formando una cellula 3n che originerà l’endosperma secondario (albume) importante nel nutrimento dell’embrione.
Durante l’evoluzione delle angiosperme si verificano modificazioni della sequenza fecondativa, ma la doppia fecondazione accomuna tutte le angiosperme e ne supporta la monofiletia.
Si ha la formazione di veri frutti, che derivano dalla trasformazione del pistillo e del ricettacolo. L’evoluzione del frutto è in stretta connessione con il tipo di dispersione adottato.
Placentazione assile.
Placentazione centrale.
Placentazione basale.
Inoltre l’ovulo può essere in diverse posizioni:
Atropo o ortotropo, cioè eretto, il tipo più primitivo.
Campilotropo, cioè orizzontale, il tipo intermedio.
Anatropo, cioè ripiegato, il tipo più comune ed evoluto.
Il gametofito femminile (sacco embrionale) non sviluppa mai archegoni. Si riduce a una struttura a 8 nuclei e 7 cellule. Degli 8 nuclei solo 3 si cellularizzano in posizione basale (cellule antipodali), altri 3 nuclei si cellularizzano nella parte apicale (cellule sinergidi) ai lati dell’oosfera. Gli ultimi 2 nuclei sono i nuclei polari liberi.
La doppia fecondazione avviene quando il tubetto pollinico penetra nelle cellule sinergidi: uno dei nuclei spermatici feconda l’oosfera, che originerà l’embrione 2n, mentre l’altro nucleo spematico si fonde con entrambi i nuclei polari formando una cellula 3n che originerà l’endosperma secondario (albume) importante nel nutrimento dell’embrione.
Durante l’evoluzione delle angiosperme si verificano modificazioni della sequenza fecondativa, ma la doppia fecondazione accomuna tutte le angiosperme e ne supporta la monofiletia.
Si ha la formazione di veri frutti, che derivano dalla trasformazione del pistillo e del ricettacolo. L’evoluzione del frutto è in stretta connessione con il tipo di dispersione adottato.
androceo : ovario
L’ovario, contenente l’ovulo con l’oosfera, durante l’evoluzione, si approfonda sempre di più nel ricettacolo (da un ovario supero a un ovario infero). Il numero di carpelli per ovario cambia durante l’evoluzione:
Ovari monocarpellari: con un solo carpello, tipico di specie più primitive.
Ovari policarpellari apocarpici: con più carpelli separati.
Ovari sincarpici più stili: con carpelli fusi.
Ovari sincarpici con uno stilo.
L’ovario sincarpico può avere un solo scomparto (uniloculare) o tre (triloculare).
L’ovulo contiene l’oosfera. L’ovulo possiede un solo tegumento o 2 ed è irrorato da un sistema conduttore (funicolo e calaza). Il funicolo e i tegumenti possono fondersi in una struttura chiamata rafe.
All’interno dell’ovulo vi è la nucella, spesso molto ridotta (ovulo tenuinucellato), ma talvolta è grossa e costituisce il perisperma (ovuli crassinucellati).
Ovari monocarpellari: con un solo carpello, tipico di specie più primitive.
Ovari policarpellari apocarpici: con più carpelli separati.
Ovari sincarpici più stili: con carpelli fusi.
Ovari sincarpici con uno stilo.
L’ovario sincarpico può avere un solo scomparto (uniloculare) o tre (triloculare).
L’ovulo contiene l’oosfera. L’ovulo possiede un solo tegumento o 2 ed è irrorato da un sistema conduttore (funicolo e calaza). Il funicolo e i tegumenti possono fondersi in una struttura chiamata rafe.
All’interno dell’ovulo vi è la nucella, spesso molto ridotta (ovulo tenuinucellato), ma talvolta è grossa e costituisce il perisperma (ovuli crassinucellati).
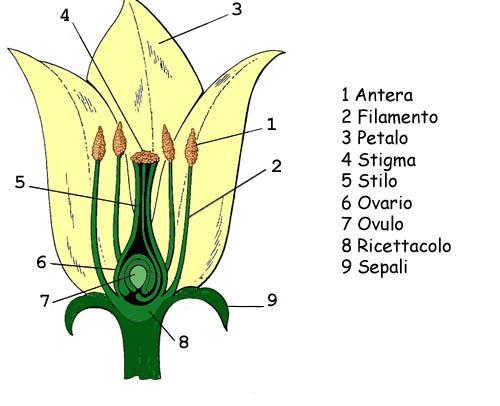
gineceo : generale

Gineceo
Il gineceo è costituito dall’insieme degli elementi sessuali femminili, i pistilli (megasporofilli). Nei progenitori delle angiosperme, il carpello è un megasporofillo richiuso e protetto da peli, ma non vi è ancora una completa fusione. Nelle angiosperme odierne, Il pistillo è costituito dall’ovario, dallo stilo e dallo stigma, posto al suo apice. Se lo stilo è assente, lo stigma è sessile.
I fiori più primitivi possono avere più pistilli posti a spirale, in seguito, nell’evoluzione, si dispongono su verticilli. Più pistilli (ginecei coricarpici o apocarpici) possono poi fondersi anche in un solo pistillo (ginecei cenocarpici o sincarpici).
Androceo
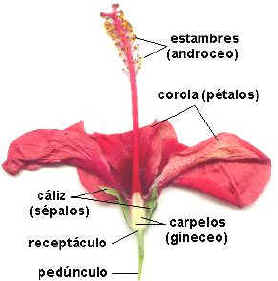
Androceo
L’androceo è composto dagli stami (microsporofilli). Essi sono costituiti da un filamento e dall’antera (dove è contenuto il polline). Nei gruppi più primitivi (Ranunculaceae) si hanno molti stami (poliandria) a disposizione spiralata, questi vengono a ridursi nei gruppi più evoluti fino ad un ciclo; anche se talvolta si può avere un aumento secondario del numero di stami (poliandria secondaria).
Gli stami delle specie più primitive sono laminari e le antere sono portate sulla lamina superiore (dorsifissi), durante l’evoluzione, subiscono riduzione progressiva (poliadi, tetradi, monadi).
Gli stami si definiscono:
Stami poliadelfi, se fusi insieme.
Stami diadelfi, se riuniti in due gruppi.
Stami monadelfi, se
L’antera è costituita da 4 secche polliniche, riunite 2 a 2, e spesso fuse in sinangi. Le antere assumono diverse posizioni (basifisse, dorsifisse, ventrifisse) e possono anche fondersi. I tessuti costituenti sono: epidermide, tessuto connettivo, tessuto meccanico (endotecio, che aiuta la fuoriuscita delle spore), tappeto plasmodiale (cellule che si disfano formando sostanze nutritive). Si definisce pollinio il contenuto dell’intera sacca pollinica; pollinario è invece il contenuto dell’intera antera.
I granuli pollinici sono microspore che germinano sullo stigma. Possiedono una parete con 1 o più pori germinativi (da cui escono i tubetti pollinici) e concamerazioni dove finiscono essudati che permettono la comunicazione con l’esterno. La parete ha una struttura complessa di scleroproteine: intina (interna) ed esina o sporopollenina (esterna) resistente all’attacco di acidi e basi forti. L’esina è dotata di un mastice pollinico, derivato dalle cellule del tappeto, che favorisce l’adesione del polline alle zampe degli insetti pronubi. Il peso dipende dal tipo di diffusione adottato.
I pollini sono anche importanti elementi diagnostici (oggetto della palinologia) e si classificano in base al numero di pori e all’ornamentazione:
Pori:
1 poro: monoporato o ulcerato.
3 pori: triporato.
Più di 3 pori: stefanoporato.
Tutto porato: pantoporato.
Solchi:
1 solco: monosolcato.
3 solchi: trisolcato.
Più di 3 solchi: stefanosolcato.
Tutto solcato: pantosolcato.
Vi può essere anche una combinazione di pori e solchi (polline colporato).
Il gametofito maschile si riduce a 2 o 3 cellule, mancano la cellula protallare e quella del peduncolo e non vi sono mai spermatozoidi. Si distinguono:
Pollini trinucleati: 2 nuclei spermatici e 1 cellula del tubetto pollinico.
Pollini binucleati: 1 nucleo spermatico e 1 cellula del tubetto pollinico.
Riproduzione grado angiosperme

Riproduzione
Il fiore possiede strutture sterili che costituiscono il perianzio (sepali e petali o tepali) e strutture fertili maschili, gli stami, e femminili, i carpelli, spesso riunite in fiori ermafroditi. Si ipotizza che il motivo della promozione dell’ermafroditismo sia stata la coevoluzione tra fiori e insetti fin dalla loro origine (entomofilia), mentre l’anemofilia, nelle angiosperme è sempre secondaria.
Le angiosperme sono le prime ad avere gli ovuli avvolti dalle foglie carpellari, evolutisi dai megasporofilli (che portavano gli ovuli). La struttura che ne deriva è detta pistillo ed è costituita dall'ovario, che contiene l'ovulo, posto sulla placenta, e dallo stilo, al cui apice è posto lo stigma sul quale germina il polline (impollinazione stigmatica). Il polline invia il tubetto pollinico attraverso il pistillo fino a raggiungere il micropilo e quindi l’oosfera.
Gli elementi sessuali nel fiore sono riuniti nell’androceo e nel gineceo.
sabato 20 dicembre 2008
tipi di infiorescenze speciali
tipi di infiorescenza
Infiorescenze speciali:
 Spighetta (Graminaceae): possiedono brattee sterili (glume) e glumette con filamenti chiamati codicule. Gli stigmi sono piumosi e sporgenti, gli stami sono lunghi e le antere rimangono fuori dalla spiga.
Spighetta (Graminaceae): possiedono brattee sterili (glume) e glumette con filamenti chiamati codicule. Gli stigmi sono piumosi e sporgenti, gli stami sono lunghi e le antere rimangono fuori dalla spiga.
 Siconio
Siconio
Ciazio: un unico ricettacolo con più fiori maschili e un fiore femminile composto da peduncolo e ovario.
Spadice
Infiorescenze speciali:
 Spighetta (Graminaceae): possiedono brattee sterili (glume) e glumette con filamenti chiamati codicule. Gli stigmi sono piumosi e sporgenti, gli stami sono lunghi e le antere rimangono fuori dalla spiga.
Spighetta (Graminaceae): possiedono brattee sterili (glume) e glumette con filamenti chiamati codicule. Gli stigmi sono piumosi e sporgenti, gli stami sono lunghi e le antere rimangono fuori dalla spiga. Siconio
SiconioCiazio: un unico ricettacolo con più fiori maschili e un fiore femminile composto da peduncolo e ovario.
Spadice
tipi di infiorescenze composte
Tipi di infiorescenza
Infiorescenze composte (o tirsoidi):
Tirso o pannocchia
Antela
Ombrella composta
Pleiocasio

Corimbo-tirso
Infiorescenze composte (o tirsoidi):
Tirso o pannocchia
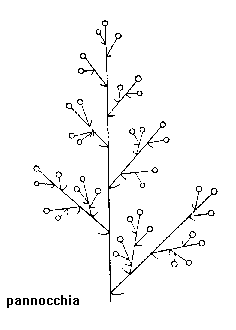
Antela

Ombrella composta

Pleiocasio

Corimbo-tirso
tipi di infiorescenze cimose

Tipi di infiorescenza
Infiorescenze cimose (o brachiali) ad accrescimento definito:
Monocasio o cima unipara (scorpioide come nelle Boraginaceae, elicoide)
Dicasio o cima ripara
Tipo di infiorescenze
Tipologia di infiorescenza:
Infiorescenze racemose (o botritiche) ad accrescimento indefinito con fioritura dal basso verso l’alto:
- Racemo

- Spiga

- Amento
- Corimbo ,Ombrella,Capolino
,Ombrella,Capolino
Infiorescenze racemose (o botritiche) ad accrescimento indefinito con fioritura dal basso verso l’alto:
- Racemo

- Spiga

- Amento

- Corimbo
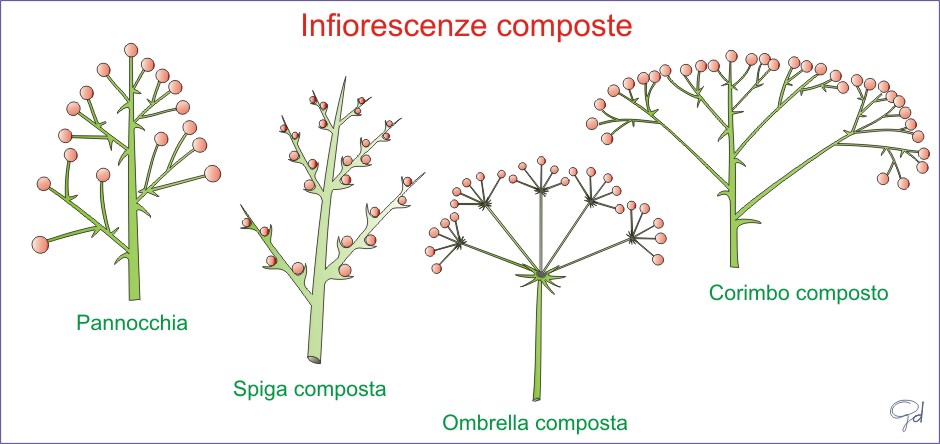 ,Ombrella,Capolino
,Ombrella,Capolino
infiorescenze nelle angiosperme
Infiorescenze
Tutti i tipi complessi di infiorescenza si evolsero da infiorescenze a pannocchia chiuse. La tendenza evolutiva è da fiori grandi a fiori piccoli, riuniti in infiorescenze che simulano un unico fiore (pseudanzio, come nelle Compositae). Alla diminuzione della dimensione dei fiori corrisponde anche l’aumento del numero degli stessi. Le foglie sono spesso inesistenti o trasformate (bratteole).
venerdì 19 dicembre 2008
Il sesso dei fiori e la determinazione della sessualità della specie nelle angiosperme
Il sesso dei fiori e la determinazione della sessualità della specie nelle angiosperme
In base al sesso dei fiori si distinguono:
In base al sesso dei fiori si distinguono:
- Fiori ermafroditi: che possiedono stami e ovari. Sono la condizione primitiva.
- Fiori unisessuali: maschili che possiedono solo stami e femminili che possiedono solo ovari. Sono la condizione derivata.
- Specie monoiche: un individuo possiede solo fiori unisessuali maschili e un altro solo fiori unisessuali femminili.
- Specie dioica: ogni individuo possiede fiori ermafroditi.
- Specie poligama: gli individui possiedono fiori sia ermafroditi che unisessuali.
Evoluzione del fiore e cambiamento della sua simmetria
Evoluzione del fiore e cambiamento della sua simmetria
Nell’evoluzione si assiste anche ad un cambiamento di simmetria del fiore:
Fiori asimmetrici primari: sono i più primitivi da cui si evolvono i fiori simmetrici.
Fiori polisimmetrici: con più di 2 piani di simmetria (multilaterali, raggiati o attinomofi).
Fiori disimmetrici: con 2 piani di simmetria (bilaterali).
Fiori monosimmetrici: con un solo piano di simmetria (dorsoventrali o zigomorfi).
Fiori asimmetrici secondari: senza piani di simmetria.
La derivazione di questi tipi di simmetria generalmente è:
Da fiori asimmetrici primari a fiori polisimmetrici.
Da fiori polisimmetrici a fiori di simmetrici.
Da fiori polisimmetrici o disimmetrici a fiori monosimmetrici.
Da fiori monosimmetrici a fiori asimmetrici secondari.
Evoluzione del fiore nelle Anguiosperme
Fiore
Nell’evoluzione del fiore si osserva una riduzione delle dimensioni ed un’oligomerizazzione dei verticilli (si riducono di numero), unita ad un accorciamento dell’asse fiorale, formando il ricettacolo. Inizialmente gli elementi fiorali si dispongono a spirale su un ricettacolo molto lungo, ma l’entomofilia ha promosso una disposizione differente del fiore. Infatti i primi insetti impollinatori erano coleotteri che, con il loro apparato masticatore, aggredivano facilmente sia il polline sia l’ovario.
Perciò l’ovario tende ad essere sempre più approfondito nel ricettacolo per avere protezione. Vi sono 2 teorie:
Teoria ricettacolare: prevede la trasformazione del ricettacolo, che da conico e allungato diviene un disco appiattito, poi incavato e infine un tubo, fondendosi con l’ovario (da ovario supero a ovario infero).
Teoria ipoanziale: prevede che le parti fiorali si fondano e chiudano l’ovario in un ipanzio.
Sulla base di questi caratteri si identificano:
Fiori aciclici primitivi (Calicanthus): privi di verticilli, con elementi fiorali posti a spirale sull’asse. Sono primitivamente asimmetrici.
Fiori emiciclici (Ranunculus, Magnolia): possiedono calice e corolla ciclici ma gli stami e l’ovario sono disposti a spirale.
Fiori ciclici (dominanti): tutti gli elementi sono posti su verticilli.
I tipi di fiore si distinguono in base al numero di verticilli che costituiscono il perianzio:
Fiori aclamidati: privi di perianzio.
Fiori aploclamidati: hanno un verticillo del perianzio.
Fiori diploclamidati: hanno due verticilli nel perianzio, si dividono in eteroclamidati, con calice (sepali) e corolla (petali) e omoioclamidati con soli tepali.
Nell’evoluzione del fiore si osserva una riduzione delle dimensioni ed un’oligomerizazzione dei verticilli (si riducono di numero), unita ad un accorciamento dell’asse fiorale, formando il ricettacolo. Inizialmente gli elementi fiorali si dispongono a spirale su un ricettacolo molto lungo, ma l’entomofilia ha promosso una disposizione differente del fiore. Infatti i primi insetti impollinatori erano coleotteri che, con il loro apparato masticatore, aggredivano facilmente sia il polline sia l’ovario.
Perciò l’ovario tende ad essere sempre più approfondito nel ricettacolo per avere protezione. Vi sono 2 teorie:
Teoria ricettacolare: prevede la trasformazione del ricettacolo, che da conico e allungato diviene un disco appiattito, poi incavato e infine un tubo, fondendosi con l’ovario (da ovario supero a ovario infero).
Teoria ipoanziale: prevede che le parti fiorali si fondano e chiudano l’ovario in un ipanzio.
Sulla base di questi caratteri si identificano:
Fiori aciclici primitivi (Calicanthus): privi di verticilli, con elementi fiorali posti a spirale sull’asse. Sono primitivamente asimmetrici.
Fiori emiciclici (Ranunculus, Magnolia): possiedono calice e corolla ciclici ma gli stami e l’ovario sono disposti a spirale.
Fiori ciclici (dominanti): tutti gli elementi sono posti su verticilli.
I tipi di fiore si distinguono in base al numero di verticilli che costituiscono il perianzio:
Fiori aclamidati: privi di perianzio.
Fiori aploclamidati: hanno un verticillo del perianzio.
Fiori diploclamidati: hanno due verticilli nel perianzio, si dividono in eteroclamidati, con calice (sepali) e corolla (petali) e omoioclamidati con soli tepali.
giovedì 18 dicembre 2008
Apparato vegetativo angiosperme
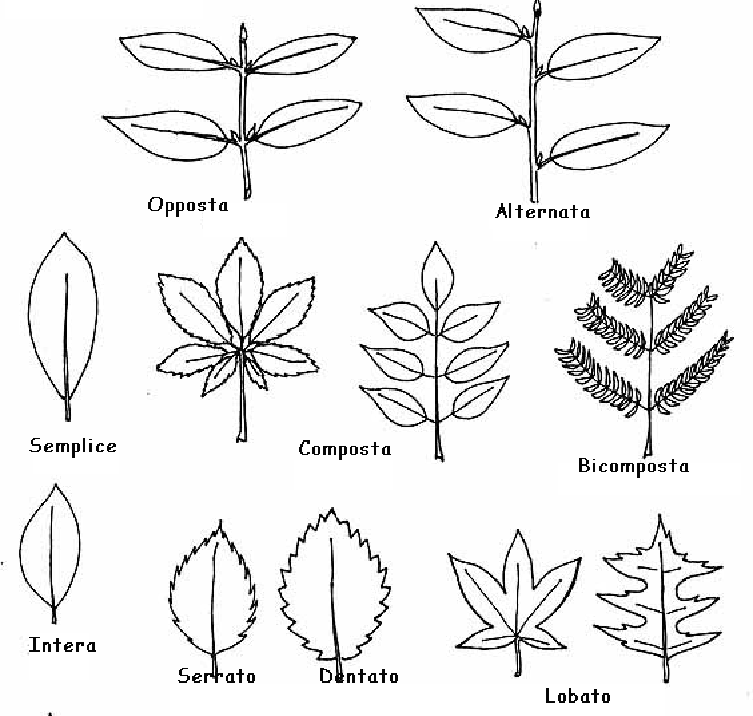
Apparato vegetativo
Presentano un portamento arboreo (generalmente simpodiale) o più comunemente erbaceo.
Il legno è eteroxilo con tracheidi e trachee con pareti più sottili. Le trachee, derivate da tracheidi scalariformi, sono un vantaggio evolutivo poiché aumentano la conduzione e permisero più vantaggi per la colonizzazione dell’ambiente terrestre, anche in climi aridi.
Il floema ha veri tubi cribrosi con specifiche cellule compagne che hanno la funzione di coordinare gli elementi dei tubi cribrosi, poiché privi di nucleo.
Le foglie semplici e intere con nervatura pennata sono la forma primitiva (secondo altri sono le foglie composte o lobate). Le nervature tendono a ramificarsi e connettersi: dalle nervature pennate derivano quelle digitate e parallele.
L’apparato radicale può essere allorizico (radice primaria e secondarie) a omorrizico secondario (solo radici secondarie).
mercoledì 17 dicembre 2008
ciclo vitale magnofolita

Ciclo vitale
L’embrione si sviluppa a spese dello sporofito (fase parassita) finché questo non viene disperso, esso è collegato da un sospensore al resto del seme.
L’embrione possiede un numero di cotiledoni da 2 (dicotiledoni) a 1 (monocotiledoni). Le specie monocotiledoni possiedono un solo cotiledone formato per fusione dei due originari. Il taxon delle Dicotiledoneae non ha più valore sistematico, in quanto parafiletico, mentre il taxon Monocotiledoneae è valido, poiché monofiletico, ma sono per lo più chiamate Liliopsida.
La plantula si nutre dei cotiledoni (come saprofita) o ancora del contenuto del seme (come parassita), dopodiché diviene verde e autotrofa.
Il ciclo vegetativo tende a divenire annuale (le piante passano l’inverno come semi) ma nelle piante perenni, generalmente legnose, si ha un periodo di stasi che coincide con la stagione avversa, generalmente l’inverno, in cui vengono perse le foglie (piante caducifoglie). Le piante che effettuano l’estivazione, perdono le foglie in estate (per esempio Euphorbia dendroides).
Il ciclo riproduttivo non coincide con il ciclo vegetativo: nelle piante perenni è ciclico ed è costituito da fioritura, fruttificazione e riposo, nelle piante annuali il seme è una fase che permette la sopravvivenza in climi aridi in modo che, quando ritorna l’acqua, viene prodotta velocemente un’altra generazione, ma se le condizioni climatiche continuano ad essere avverse, il seme rimane in stato di riposo.
Dopo aver prodotto la nuova generazione la pianta annuale, muore.
Caratteri che supportano la monofiletia delle angiosperme
Caratteri che supportano la monofiletia delle angiosperme
L’insieme dei caratteri che portano a supporre un’origine monofiletica delle angiosperme da un comune gruppo ancestrale di gimnosperme sono molti.
Non vi è discontinuità morfologica evidente rispetto alle altre spermatofite.
Non vi è discontinuità morfologica evidente all’interno delle angiosperme stesso.
Possiedono molti caratteri apomorfi, comuni a tutte le angiosperme:
Tubi cribrosi e cellule compagne da una cellula iniziale comune.
Fiori primitivi con stami inseriti inferiormente ai carpelli.
Stami con 2 gruppi di sacche polliniche ed 1 endotecio.
Carpelli chiuse.
Gametofito maschile con 3 cellule.
Gametofito femminile con struttura a 8 nuclei e 7 cellule (primitivamente).
Doppia fecondazione.
Endosperma secondario triploide.
martedì 16 dicembre 2008
GRADO ANGIOSPERME Sudivisio Magnoliophytina

Sudivisio Magnoliophytina
Compaiono nel Giurassico ed evolvono e diversificano nel Cretaceo, per arrivare nel Quaternario dove sono il gruppo di spermatofite con maggior numero di specie. Non si conosce ancora il loro gruppo primitivo, da cui sono derivate, poiché vi sono lacune nei fossili. L'ipotesi oggi più accettata è che le magnoliofitine siano derivate dalle Lyginopteridopsida (Pteridospermae) o dalle Bennettitopsida.
lunedì 15 dicembre 2008
Subclassis Ephedridae
Subclassis Ephedridae
Ordo Ephedrales
Ephedra
Vivono in zone temperato-calde. Piante arbustive, con fusti verdi molto ramificati, le foglie sono squamiformi opposte o verticillate.
Sono per lo più dioiche, ma vi sono anche specie moniche. I fiori sono posti all'ascella di brattee decussate o in cima ai rami: i fiori maschili hanno uno stame e un perianzio bilobato, i fiori femminili hanno un ovulo con un micropilo rialzato e un perianzio bilobato. I fiori possono anche essere riuniti in infiorescenze ermafrodite. L'impollinazione è entomofila con sirfidi e apidi pronubi.
Si osserva doppia fecondazione, tipica delle angiosperme, fatto sul quale alcuni autori identificano un gruppo monofiletico con le angiosperme: i due spermazi contenuti nel gametofito maschile hanno destini differenti. Uno feconda l'oosfera e originerà perciò l'embrione, l'altro si fonderà con i nuclei vegetativi a dare un tessuto trofico, l'endosperma secondario, che nutrirà l'embrione. Questo è un vantaggio evolutivo, poiché permette un risparmio energetico nel nutrimento di un embrione.
Ordo Ephedrales
Ephedra
Vivono in zone temperato-calde. Piante arbustive, con fusti verdi molto ramificati, le foglie sono squamiformi opposte o verticillate.
Sono per lo più dioiche, ma vi sono anche specie moniche. I fiori sono posti all'ascella di brattee decussate o in cima ai rami: i fiori maschili hanno uno stame e un perianzio bilobato, i fiori femminili hanno un ovulo con un micropilo rialzato e un perianzio bilobato. I fiori possono anche essere riuniti in infiorescenze ermafrodite. L'impollinazione è entomofila con sirfidi e apidi pronubi.
Si osserva doppia fecondazione, tipica delle angiosperme, fatto sul quale alcuni autori identificano un gruppo monofiletico con le angiosperme: i due spermazi contenuti nel gametofito maschile hanno destini differenti. Uno feconda l'oosfera e originerà perciò l'embrione, l'altro si fonderà con i nuclei vegetativi a dare un tessuto trofico, l'endosperma secondario, che nutrirà l'embrione. Questo è un vantaggio evolutivo, poiché permette un risparmio energetico nel nutrimento di un embrione.
Subclassis Gnetidae
Subclassis Gnetidae
Ordo Gnetales
Gnetum
Sono tropicali e sub-tropicali. Per la maggior parte sono lianose, ma vi sono anche arboree e arbustive. Le foglie sono ellittiche retinervie.
Vi sono specie moniche e dioiche. I fiori sono unisessuali e posti all'ascella di squame circolari concresciute in infiorescenze spiciformi: i fiori maschili sono costituiti da un unico stame circondato da un'unica brattea perinaziale, i fiori femminili possiedono un ovulo, con due tegumenti di cui quello interno allungato in un tubo per sollevare il micropilo, e un perianzio. L'impollinazione è entomofila.
L'endosperma è sia cellularizzato che liquido, perciò tende ad uscire e formare un liquido di impollinazione.
domenica 14 dicembre 2008
Subclassis Welwitschidae
Subclassis Welwitschidae
Ordo Welwitschiales
Welwitschia mirabilis
Vive nel deserto della Namibia. Possiede un fusto conico affiorante alla superficie del suolo ed originato dall'ipocotile, da cui dipartono due sole foglie, opposte, che grazie ad un meristema basale hanno accrescimento indefinito (fino a 3 m), sfrangiandosi in fondo. Le foglie fanno da imbuto e condensano la nebbia in acqua liquida, che viene raccolta dalla coppa del fusto dove sporgono le infiorescenze. La radice è fittonosa e si approfonda per cercare le falde acquifere.
La specie è dioica e le strutture riproduttive sono riunite in strobili. Il fiore maschile ha stami e brattee perianziali, il fiore femminile ha un ovulo con tegumento spesso e brattee perianziali. Da notare che anche il fiore maschile porta un ovario rudimentale, perciò è probabile che in origine i loro antenati fossero ermafroditi, in cui sia avvenuta una separazione dei sessi. L'impollinazione è entomofila ma ancora micropilare, perciò il micropilo è posto su un filamento per aumentare la probabilità di impollinazione.
Ordo Welwitschiales
Welwitschia mirabilis
Vive nel deserto della Namibia. Possiede un fusto conico affiorante alla superficie del suolo ed originato dall'ipocotile, da cui dipartono due sole foglie, opposte, che grazie ad un meristema basale hanno accrescimento indefinito (fino a 3 m), sfrangiandosi in fondo. Le foglie fanno da imbuto e condensano la nebbia in acqua liquida, che viene raccolta dalla coppa del fusto dove sporgono le infiorescenze. La radice è fittonosa e si approfonda per cercare le falde acquifere.
La specie è dioica e le strutture riproduttive sono riunite in strobili. Il fiore maschile ha stami e brattee perianziali, il fiore femminile ha un ovulo con tegumento spesso e brattee perianziali. Da notare che anche il fiore maschile porta un ovario rudimentale, perciò è probabile che in origine i loro antenati fossero ermafroditi, in cui sia avvenuta una separazione dei sessi. L'impollinazione è entomofila ma ancora micropilare, perciò il micropilo è posto su un filamento per aumentare la probabilità di impollinazione.
sabato 13 dicembre 2008
Classis Gnetopsida

Classis Gnetopsida (= Clamidospermae; Subdivisio Clamidospermophytina)
Alcuni autori riuniscono le Bennettitopsida e le Gnetopsida nella divisione delle Anthophyta, poiché considerano le bennettitopside antenate delle gnetopside in quanto le prime possiedono un fiore meno evoluto delle seconde. Ma i loro rapporti non sono così diretti e perciò l'ipotesi è decaduta.
Le gnetopside sono un gruppo filogeneticamente dubbio. Vi sono contenuti 3 generi:
Welwitschia, del deserto della Namibia (1 specie).
Gnetum, con specie in India, Cina, Guinea, Australia e Brasile.
Ephedra, dell’America meridionale, ma si trova anche in Italia: in Trentino e in Sardegna.
Questi 3 generi sono posti in sottoclassi differenti, poiché il loro progenitore comune non è vicino, dato che condividono pochi caratteri:
Saldatura parziale della foglia carpellare (quindi un ovario ancora più protetto).
Fiori muniti di antofilli perianziali.
Legno eteroxilo (tracheidi e trachee).
Riduzione progressiva e costante del gametofito.
Diffusa poliembrionia.
Embrione con 2 cotiledoni.
Inoltre alcuni di questi caratteri sono oggi messi in dubbio: le trachee che danno il legno eteroxilo sono in realtà derivate dalla fusione e allungamento di tracheidi con punteggiature areolate, in pseudotrachee. Anche il fiore, prima considerato omologo a quello delle angiosperme, oggi è considerato analogo.
La paleontologia non ha aiutato a chiarire la loro posizione, poiché sono un gruppo originatosi alla fine del Terziario, forse originatosi dalle bennettitopside. Probabilmente il taxon Gnetopsida è un gruppo di comodo.
Subclassis Pentoxylidae
Subclassis Pentoxylidae
Ordo Pentoxylales
Possiedono elementi meno evoluti rispetto alle bennettitali, perciò alcuni autori le classificano a parte.
Il fusto presentava una polistele. Possedevano foglie linguiformi penninervie. Le squame interseminali erano assenti. I semi maturi erano disposti in strobili simili ad angiosperme primitive (Pandanus).
Ordo Pentoxylales
Possiedono elementi meno evoluti rispetto alle bennettitali, perciò alcuni autori le classificano a parte.
Il fusto presentava una polistele. Possedevano foglie linguiformi penninervie. Le squame interseminali erano assenti. I semi maturi erano disposti in strobili simili ad angiosperme primitive (Pandanus).
venerdì 12 dicembre 2008
Classis Bennettitopsida +
Classis Bennettitopsida +
Subclassis Bennettitidae
Ordo Bennettitales
Sono tutte estinte. L'aspetto generale era simile alle cicadopside, infatti potevano essere pachicauli o leptocauli con ramificazione simpodiale e possedevano foglie pennate o interne.
Ma la somiglianza era solo superficiale, poiché le bennettitali presentavano veri fiori, cioè stami maschili (microsporofilli) e ovuli (megasporofilli) avvolti da strutture sterili simili al perianzio. Gli ovuli erano unici per ogni fiore e peduncolati, poggiando direttamente sull'asse fiorale. Vi erano squame interseminali disposte a spirale. Il tegumento degli ovuli (forse era doppio in origine) sporgeva dalle squame interseminali. Gli stami erano in parte pennati, lobati o semplificati. Le sacche polliniche formavano sinangi marginali o erano immerse nella pagina superiore dello stame. Secondo alcuni autori gli stami si staccavano dalla pianta e si aprivano a terra attirando insetti pronubi.
Alcune specie erano già ermafrodite (Williamsonia aveva ancora fiori unisessuali). Si è ipotizzato che fossero già proterandre, cioè che per evitare l'autofecondazione si sviluppassero prima gli stami, che potevano disperdere il polline per fecondare altre piante che avevano ovuli già maturi. Questa serie di somiglianze ha fatto ipotizzare che le bennettitali potessero essere antenate delle angiosperme, ma oggi si tende a considerarle convergenze evolutive.
L'impollinazione era entomofila (probabilmente ad opera di coleotteri). Probabilmente possedevano ancora spermatozoidi.
Oltre al fiore era presente anche un frutto analogo che racchiudeva il seme in strati carnosi sterili. L'embrione possedeva 2 cotiledoni.
giovedì 11 dicembre 2008
Familia Zamiaceae

Familia Zamiaceae
Zamia
Possiede struttura simpodiale. Sia i megasporofilli che i microsporofilli sono riuniti in strobili. I megasporofilli portano 2 ovuli. Gli spermatozoidi hanno una lunghezza record: 1/2 mm.
mercoledì 10 dicembre 2008
Cycas revoluta

Cycas revoluta
E' la specie più comune, molto usata come pianta ornamentale. Possiede struttura monopodiale, anche se le ramificazioni sono rare e solo negli indivisi più vecchi. Possiede foglie fertili e sterili poste ad anello. Le radici si intrecciano fittamente e sono in simbiosi con alghe azzurre azotofissatrici (Anabaena).
Si distingue soprattutto per i megasporofilli non riuniti in strobili e microsporofilli riuniti in strobili. I megasporofilli portano più di 2 ovuli, questi sono molto grandi e si accrescono anche se non fecondati (è un carattere primitivo).
Nelle piante maschili, dopo che si forma lo strobilo, si ha un blocco nell'accrescimento, mentre nelle piante femminili si ha accrescimento indefinito, alternando tra foglie sterili e fertili.
martedì 9 dicembre 2008
classis Cycadopsida

classis Cycadopsida
Ordo Cycadales
Familia Cycadaceae
La classe delle cicadopside ebbe il suo massimo sviluppo nel Giurassico. Oggi sopravvivono solo poche specie, fossili viventi, appartenenti alla flora dell'America Centrale e dell'Australia.
Sono generalmente pachicauli e la loro morfologia esterna è molto simile alle palme, poiché possiedono uno stipite colonnare con ciuffi di foglie apicali disposte a spirale. Le foglie sono grandi, pennate o bipennate, ed hanno crescita apicale continua. Le nervature sono pennate e spesso dicotomiche aperte. In sezione è ben visibile il tessuto di trasfusione, costituito da cellule simili a tracheidi che collaborano al trasporto dei liquidi dalle nervature fogliari al mesofilli. Gli stomi sono aplocheilici, perciò le cellule di guardia non sono omologhe alle cellule annesse.
Gli sporotrofilli presentano una riduzione della parte sterile, in una sequenza progressiva nelle differenti specie. Sono piante dioiche: gli individui maschili possiedono gruppi di sacche polliniche ventrali sui microsporofilli, le piante femminili possiedono ovuli su megasporofilli poco differenziati in strobili o in strobili molto semplici. L'ovulo possiede 2 strutture: una camera pollinica e una camera archegoniale.
L'impollinazione è anemofila (tranne in Encephalarthos dove è entomofila per mezzo di coleotteri) e tra impollinazione e fecondazione intercorrono alcuni mesi. Il polline viene catturato con una goccia di impollinazione, in seguito raggiunge la camera pollinica dove germina e origina un tubetto pollinico austoriale che permette al gametofito maschile di lisare la nucella per raggiungere la camera archegoniale, dove si trovano gli archegoni con le oosfere. Gli spermatozoidi sono pluriflagellati.
Il seme possiede sarcotesta e sclerotesta ben sviluppate.
Molte specie delle cicadacee sono tossiche e cancerogene. I semi vengono trattati per utilizzarne l'amido.
lunedì 8 dicembre 2008
Familia Caytoniaceae
Ordo Caytoniales
Familia Caytoniaceae
Caytonia
Le foglie erano palmate o pennate e retinervie. Aveva una struttura particolare dei megasporofilli: erano pennati, portavano molti ovuli e tendevano a richiudersi sopra di essi. Si è avanzata l'ipotesi che i fiori delle angiosperme siano derivati da questa struttura, ma oggi non è un'ipotesi valida.
Gli stami avevano molti sinangi di 4 sacche polliniche e i granuli pollinici possedevano sacche aerifere, perciò avevano impollinazione anemofila. Vi sono tracce di una goccia di impollinazione per trattenere il polline.
Familia Caytoniaceae
Caytonia
Le foglie erano palmate o pennate e retinervie. Aveva una struttura particolare dei megasporofilli: erano pennati, portavano molti ovuli e tendevano a richiudersi sopra di essi. Si è avanzata l'ipotesi che i fiori delle angiosperme siano derivati da questa struttura, ma oggi non è un'ipotesi valida.
Gli stami avevano molti sinangi di 4 sacche polliniche e i granuli pollinici possedevano sacche aerifere, perciò avevano impollinazione anemofila. Vi sono tracce di una goccia di impollinazione per trattenere il polline.
domenica 7 dicembre 2008
Familia Glossopteridaceae
Familia Glossopteridaceae
Avevano un fusto con eustele o polistele. Possedevano gonofilli da cui è possibile sia originato il fiore. Erano costituiti da foglie ovali o laminari con nervature reticolate chiuse e con apici fertili. Potevano essere sia monoiche che dioiche.
sabato 6 dicembre 2008
Ordo Lyginopteridales
Ordo Lyginopteridales

Alberi o liane con grandi foglie pennate (Lyginopteridaceae, Medullosaceae) o piante erbacee rizomatose con foglie allungate linguiformi e con nervature laterali anastomizzate. Il fusto presentava una protostele o una eustele con un mantello di legno secondario (Lyginopteridaceae) o polistele (Medullosaceae).
Il tegumento del seme era suddiviso in 3 strati evidenti e fasci vascolari nel tegumento e nella nucella (Lyginopteridaceae, Medullosaceae). La cupola era saldata al tegumento del seme (Medullosaceae) o libera (Lyginopteridaceae).

Alberi o liane con grandi foglie pennate (Lyginopteridaceae, Medullosaceae) o piante erbacee rizomatose con foglie allungate linguiformi e con nervature laterali anastomizzate. Il fusto presentava una protostele o una eustele con un mantello di legno secondario (Lyginopteridaceae) o polistele (Medullosaceae).
Il tegumento del seme era suddiviso in 3 strati evidenti e fasci vascolari nel tegumento e nella nucella (Lyginopteridaceae, Medullosaceae). La cupola era saldata al tegumento del seme (Medullosaceae) o libera (Lyginopteridaceae).
venerdì 5 dicembre 2008
Classis Lyginopteridopsida (= Pteridospermae) +
Classis Lyginopteridopsida (= Pteridospermae) +
Sono anche chiamate felci a seme. Comparvero nel Devoniano superiore e si estinsero all'inizio del Cretaceo.
Avevano portamento arboreo con fusto a stipite colonnare e fronde ramificate, viceversa potevano essere a fusto lianoso appoggiandosi ad altre piante.
Si osserva una tendenza da trofosporofilli e sporofilli con una riduzione della parte sterile e del numero di ovuli, ma non formano mai fiori. Ciò ha fatto ipotizzare la derivazione dell'ovario delle gimnosperme da questo tipo di sporofilli perciò erano considerate di passaggio tra le progimnosperme e le gimnosperme. L'ipotesi oggi più accettata è che abbiano originato tutte le altre cicadofitine e le angiosperme.
Gli ovuli avevano più tegumenti e si individuavano:
Cupola, che avvolgeva tutto l'ovulo.
Nucella, spesso fusa con il primo tegumento.
Camera pollinica, che accoglieva gli spermatozoidi ancora flagellati.
Micropilo è ampio e con lobi ramificati
Si osserva anche il passaggio da telomi terminali, intorno all'ovulo, fino alla fusione di questi nella cupola. La cupola avvolgeva un unico ovulo o numerosi. In alcune specie i tegumenti dell'ovulo si prolungavano analogamente a stili e stimmi delle angiosperme.
Dalla cellula madre delle spore solo una portava a compimento il suo sviluppo, le altre rimanevano all'apice della megaspora. Il seme possedeva una sarcotesta ed una sclerotesta.

Sono anche chiamate felci a seme. Comparvero nel Devoniano superiore e si estinsero all'inizio del Cretaceo.
Avevano portamento arboreo con fusto a stipite colonnare e fronde ramificate, viceversa potevano essere a fusto lianoso appoggiandosi ad altre piante.
Si osserva una tendenza da trofosporofilli e sporofilli con una riduzione della parte sterile e del numero di ovuli, ma non formano mai fiori. Ciò ha fatto ipotizzare la derivazione dell'ovario delle gimnosperme da questo tipo di sporofilli perciò erano considerate di passaggio tra le progimnosperme e le gimnosperme. L'ipotesi oggi più accettata è che abbiano originato tutte le altre cicadofitine e le angiosperme.
Gli ovuli avevano più tegumenti e si individuavano:
Cupola, che avvolgeva tutto l'ovulo.
Nucella, spesso fusa con il primo tegumento.
Camera pollinica, che accoglieva gli spermatozoidi ancora flagellati.
Micropilo è ampio e con lobi ramificati
Si osserva anche il passaggio da telomi terminali, intorno all'ovulo, fino alla fusione di questi nella cupola. La cupola avvolgeva un unico ovulo o numerosi. In alcune specie i tegumenti dell'ovulo si prolungavano analogamente a stili e stimmi delle angiosperme.
Dalla cellula madre delle spore solo una portava a compimento il suo sviluppo, le altre rimanevano all'apice della megaspora. Il seme possedeva una sarcotesta ed una sclerotesta.
Subdivisio Cycadophytina
Subdivisio Cycadophytina

Sono sia pachicauli che leptocauli e presentano ramificazione dicotoma, monopodiale o simpodiale. Le tracheidi dello xilema sono scalariformi o con punteggiature areolate. Le foglie sono pluripennate oppure indivise nastriformi o squamiformi. Le nervature sono dicotome, pennate o reticolate.
Gli organi sessuali possono trovarsi associati a trofofilli (trofosporofilli) oppure in sporofilli divisi in maschili e femminili. In alcune specie si evolve anche un fiore: sono sporofilli riuniti su brachiblasti a crescita limitata.
Gli stami hanno diversi gruppi di sacche polliniche fuse (sinangi). I megasporofilli portano parecchi ovuli almeno nelle specie più primitive.
L'impollinazione in genere è anemofila, ma talora è zoofila.
mercoledì 3 dicembre 2008
Subclassis Taxidae
Subclassis Taxidae
Familia Taxaceae
Taxus baccata
Possiede habitus arboreo o arbustivo ed è molto utilizzato come pianta ornamentale per la sua folta chioma scura. Non possiede i tipici canali resiniferi delle Pinidae.
Contiene un veleno mortale e l'unica parte non velenosa è un cercine rosso che circonda il seme, l'arillo. Questo permette la diffusione endozoocora poiché il seme possiede un rivestimento duro, la sclerotesta, e non viene digerito.
I fiori femminili sono pseudoterminali con alla base alcune coppie di squame e 1 solo ovulo apicale eretto. Gli stami sono peltati e portano diverse sacche polliniche. L'embrione possiede due cotilendoni.
Familia Taxaceae
Taxus baccata
Possiede habitus arboreo o arbustivo ed è molto utilizzato come pianta ornamentale per la sua folta chioma scura. Non possiede i tipici canali resiniferi delle Pinidae.

Contiene un veleno mortale e l'unica parte non velenosa è un cercine rosso che circonda il seme, l'arillo. Questo permette la diffusione endozoocora poiché il seme possiede un rivestimento duro, la sclerotesta, e non viene digerito.
I fiori femminili sono pseudoterminali con alla base alcune coppie di squame e 1 solo ovulo apicale eretto. Gli stami sono peltati e portano diverse sacche polliniche. L'embrione possiede due cotilendoni.
martedì 2 dicembre 2008
Taxodium disticum

Taxodium disticum
E' tra le poche caducifoglie originarie dell'America Sud Orientale. Vive in zone paludose e possiede perciò radici aeree respiratorie con all'apice pneumatofore.
Sequoia sempervirens

Sequoia sempervirens
Raggiunge fino a 100 m di altezza ma ha un diametro relativamente piccolo. Vive generalemente 1000 anni. La sequoia più alta conosciuta è il General Sherman.
Familia Taxodiaceae

Sono tra gli alberi di maggiori dimensioni. Sono estranee alla nostra flora, le specie autoctone sono note allo stato fossile e quelle viventi derivano da coltivazione. Attualmente sono autoctone dell'America con il genere Sequoia e della Cina con il genere Metasequoia.
Grazie alla loro longevità sono spesso usate in dendrocronologia.
lunedì 1 dicembre 2008
Familia Cupressaceae
 Comprende i generi Cupressus e Juniperus. Le foglie sono aghiformi o squamiformi, tuttavia nelle specie con foglie squamiformi, le giovani foglie, attraversano uno stadio aghiforme (ricapitolazione).
Comprende i generi Cupressus e Juniperus. Le foglie sono aghiformi o squamiformi, tuttavia nelle specie con foglie squamiformi, le giovani foglie, attraversano uno stadio aghiforme (ricapitolazione).
Possiedono un numero di ovuli superiore a 2 per ogni squama ovulifera, poiché queste concrescono e si fondono. I galbuli sono gli strobili maturi del genere Juniperus e sono commestibili, in modo da permettere la dispersione endozoocora.
In Cupressus vi sono foglioline embricate e i galbuli maturi sono lignificati.
domenica 30 novembre 2008
Pinus cembra

Pinus cembra
Il pino cembro è l'unico pino autoctono italiano che invece di avere 2 aghi per brachiblasto ne ha 5.
sabato 29 novembre 2008

venerdì 28 novembre 2008
Pinus sylvatica

Pinus sylvatica
Possiede scorza rossastra, aghi spiralati e pigne piccole. E' autoctono dell'Italia.
giovedì 27 novembre 2008
Pinus laricio

Pinus laricio
Il pino nero è originario delle montagne del Sud Italia. Possiede aghi robusti simili a quelli di P. pinaster ma molto più scuri. La scorza invece è chiara ma spesso annerita da incendi.
mercoledì 26 novembre 2008
Pinus aleppensis

Pinus aleppensis
Il pino d'aleppo cresce spontaneo in zone costiere e rupestri, ma è stato anche introdotto artificialmente. La chioma è scarmigliata, costituita da aghi sottili e leggeri. La scorza è grigio chiaro. Le pigne raggiungono gli 8 cm di lunghezza e sono peduncolate. Il loro tempo di maturazione è lungo: all'inizio presentano brattee rosse, dopo la fecondazione sono verdi e si accrescono. Su una stessa pianta vi possono essere quindi tutti gli stadi di maturazione.
martedì 25 novembre 2008
Pinus pinaster

Pinus pinaster
Il pino marittimo possiede aghi lunghi e robusti e pigne allungate. Il luogo di origine è stato individuato nella Francia meridionale. Le popolazioni attuali sono quasi tutte introdotte e sono favorite dal fuoco: il calore apre le pigne e i pinoli sopravvivono grazie al loro involucro coriaceo. Le plantule sono eliofile, perciò avendo molto spazio aperto crescono in fretta.
Iscriviti a:
Post (Atom)



{kind=link}